Experiments in the Morrissette laboratory are directed at understanding how microtubules function in Apicomplexan parasites. The radically different organization, regulation and use of microtubules in these protozoa represent intriguing departures from our understanding of microtubules in model organisms. Moreover, these distinct properties can be exploited to develop novel anti-parasitic therapies. To establish basic principles, we work with Toxoplasma gondii because of its relative ease of manipulation. Ultimately we will extend our analysis to other apicomplexans, particularly Plasmodium spp.
Microtubules, tubulin & tubulin-binding drugs:
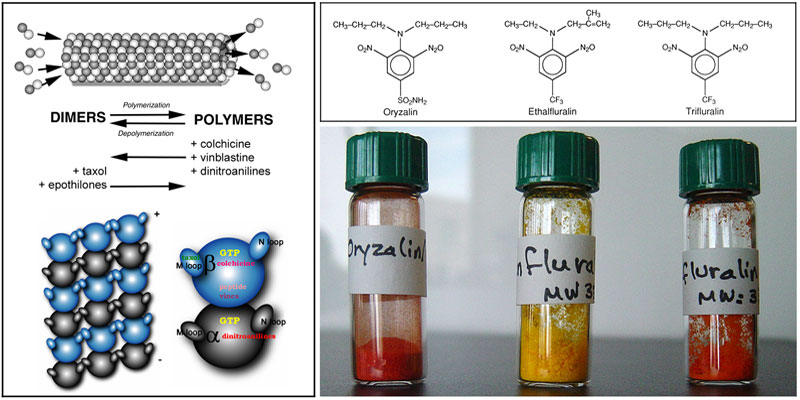
Microtubules are polymers constructed from a-b-tubulin heterodimers (Figure 7). These structures are rapidly assembled and disassembled to create essential components of eukaryotic cells, such as spindles and flagella. The dynamic nature of microtubules makes them susceptible to pharmacological agents. Microtubule-disrupting and microtubule-stabilizing drugs have provided great insight into tubulin and microtubule function; they also have tremendous practical use. Compounds that perturb microtubule dynamics are currently some of the most effective drugs to treat medical conditions including cancer, gout and helminth infection.
 |
|
| Figure 7: Inside eukaryotic cells, tubulin is in equilibrium between the disassembled dimer “subunit” form and the assembled microtubule “polymer” form. Microtubules have a substructure called the protofilament which is formed by the head to tail association of dimers. Under most circumstances, 13 protofilaments associate to form microtubules. Both a and b tubulin have M and N loops, regions that coordinate the lateral interactions between protofilaments and which are critically important to regulating the microtubule lattice stability. Diverse pharmacological agents shift this equilibrium toward one or the other extreme. Taxol and the epothilones stabilize microtubules. Colchicine, vinblastine and the dinitroanilines disassemble microtubules. Notably, the dinitroanilines are the first category of compounds that bind to a-tubulin; all previously characterized drugs bind to b-tubulin. Many dinitroaniline compounds have been used for many years are as commercial herbicides. Oryzalin (Surflan), ethalfluralin (Sonalan) and trifluralin (Treflan) are used as pre-emergence herbicides to kill grasses and broadleaf weeds. We most commonly use oryzalin in our Toxoplasma experiments, but ethalfluralin and trifluralin also disrupt parasite microtubules in a similar fashion. | |
Toxoplasma microtubule populations:
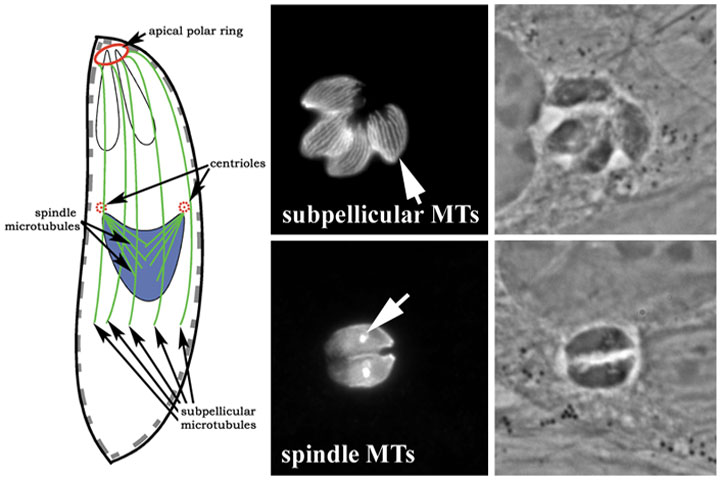
The haploid asexual forms of Toxoplasma (tachyzoites, bradyzoites and sporozoites) have two populations of microtubules: spindle microtubules and subpellicular microtubules (Figure 8). Spindle microtubules (~1-2m long) function to form an intra-nuclear spindle (a closed mitosis) to coordinate chromosome segregation. The spindle microtubules originate in a dense plaque structure that is embedded in the nuclear membrane adjacent to cytoplasmic centrioles. The characteristic crescent shape of Toxoplasma is maintained by an interaction between the pellicle and the underlying twenty-two subpellicular microtubules. The pellicle is composed of the plasma membrane and the closely apposed inner membrane complex that is comprised of flattened vesicles. The subpellicular microtubules (~5m long) have a characteristic organization and length and are nucleated from the apical polar ring, a microtubule-organizing center (MTOC) that is unique to the Apicomplexa. These microtubules are also required to maintain apical polarity. Both populations are critically important to parasite survival and replication. Although extracellular parasites are refractory to the effects of microtubule-disrupting drugs, during intracellular growth parasite microtubules are dynamic and are highly sensitive to disruption.
 |
||
| Figure 8: Toxoplasma tachyzoites have two populations of microtubules: spindle microtubules and subpellicular microtubules. Spindle microtubules form an intra-nuclear spindle to coordinate chromosome segregation during mitosis. The spindle microtubules originate in a dense plaque structure that is embedded in the nuclear membrane adjacent to cytoplasmic centrioles. The characteristic crescent shape and apical polarity of Toxoplasma is maintained by an interaction between the pellicle and the underlying twenty-two subpellicular microtubules. These microtubules are nucleated by the apical polar ring. | ||
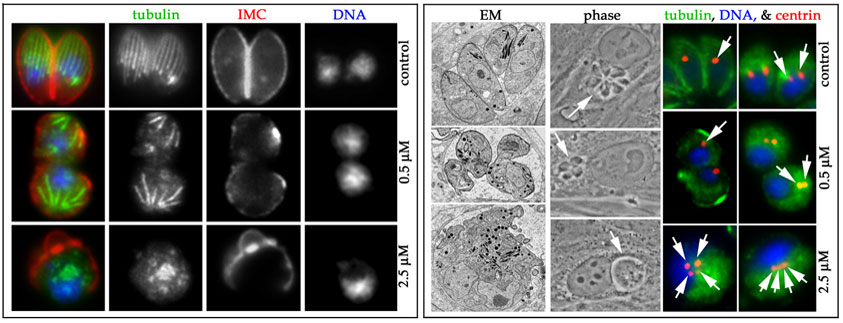
In cell biological studies, we have established that the two populations of microtubules in Toxoplasma have distinct susceptibilities to oryzalin and other dinitroanilines (Figure 9). Each of these microtubule populations is individually nucleated by a distinct microtubule-organizing center (MTOC): the apical polar ring for the subpellicular microtubules and nuclear plaques/centrioles for the spindle microtubules. When replicating Toxoplasma are treated with 0.5 mM oryzalin, they retain the capacity to form a spindle and undergo nuclear division. Although these parasites can complete budding, their subpellicular microtubules are shortened or absent and they lose the ability to reinvade host cells. Both spindle and subpellicular microtubules are disrupted in 2.5 mM oryzalin. Under these conditions, parasites grow in size and replicate their DNA but are incapable of nuclear division. We believe the differential susceptibility of subpellicular and spindle microtubule is primarily due to the required length that each of these populations must attain to be functional. Normal subpellicular microtubules are ~5m in length while spindle microtubules are ~1m. In 0.5 mM oryzalin, parasites can build short microtubules. These are sufficient to function in spindles but not as subpellicular microtubules.
 |
|||
|
Figure 9: Toxoplasma subpellicular microtubules and spindle microtubules are differentially sensitive to oryzalin disruption. In the absence of oryzalin, Toxoplasma have an elongated cell shape as visualized by immunofluorescence, phase-contrast and electron microscopy. In 0.5 mM oryzalin, the subpellicular microtubules are shortened and consequently, parasites are round. Nuclear division proceeds with correct segregation of the centrioles and budding of daughter parasites in 0.5 mM oryzalin. However, in 2.5 mM oryzalin both spindle and subpellicular microtubules are disrupted, nuclear division and budding cease and centrioles continue to duplicate unchecked. (Left box) Triple immunofluorescence of microtubules (green), inner membrane complex (red) and DNA (blue) in control cells, samples treated with 0.5 mM oryzalin or samples treated with 2.5 mM oryzalin. Without drug, the subpellicular microtubules extend along two-thirds of the length of crescent-shaped parasites. In 0.5 mM oryzalin, the subpellicular microtubules are shortened although nuclear division continues with nuclear segregation. In 2.5 mM oryzalin, all microtubules are disrupted and the parasite grows as a large round blob. (Right box) Electron microscopy, phase-constrast microscopy and immunofluorescence of parasites. The triple immunofluorescence images show tubulin (green), centrin (red) and DNA (blue). Centrioles (labeled with centrin) appear as a single (non-replicated) spot or two (replicated) spots in untreated parasites. |
|||
Dinitroaniline action on Toxoplasma:
One primary goal of our research is to understand the basis of dinitroaniline action on Toxoplasma. Specifically, understanding why these compounds are selectively active against plant and protozoan tubulins will be important to design therapeutic agents that selectively disrupt parasite, but not host, microtubules. In addition, by understanding the diverse mechanisms of parasite resistance to dinitroanilines, we hope to identify the genes for proteins that regulate microtubule stability in the Apicomplexa. In the unicellular alga Chlamydomonas, many mutations that confer dinitroaniline resistance map to microtubule regulatory proteins, rather than to tubulin.
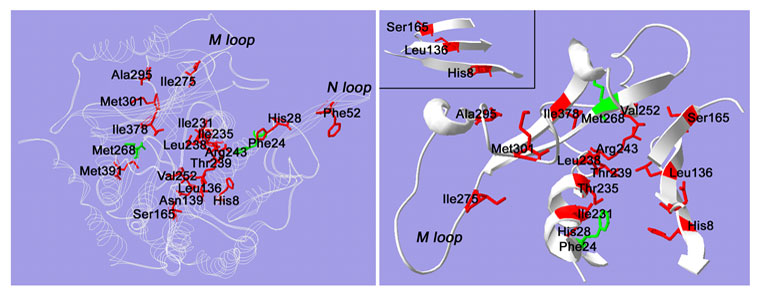
Our recent studies have demonstrated that low-level resistance to the dinitroaniline oryzalin is invariably associated with point mutations to a-tubulin. The point mutations cluster into specific regions of the a-tubulin structure, namely a core region and the M and N loops (Figure 10). These point mutations are sufficient to confer oryzalin resistance when introduced into wild-type (sensitive) parasites.
 |
||
| Figure 10: The amino acid sequence of Toxoplasma a-tubulin was fit to the structure of bovine a-tubulin using Swiss-Model automated comparative protein modeling. The top panel shows residues that are mutated in oryzalin-resistant Toxoplasma. The plant mutations at residues 24 and 268 are colored green. The mutations fall into three areas: the M loop, the N loop and a core region. The bottom panel shows cut-away views of some of the core residues that are mutated to confer oryzalin resistance. Many residues are in striking proximity and show specific orientations. For example, residues Ile231, Ile235 and Thr239 all occur on the same face of a-helix 7. Leu238, Arg243 and Val252 cluster in the same general area. The inset shows a b-sheet formed by strands 1, 4 and 5. The mutated residues His8, Leu136 and Ser165 are in a linear pattern across the b-sheet. | ||
 |
|||
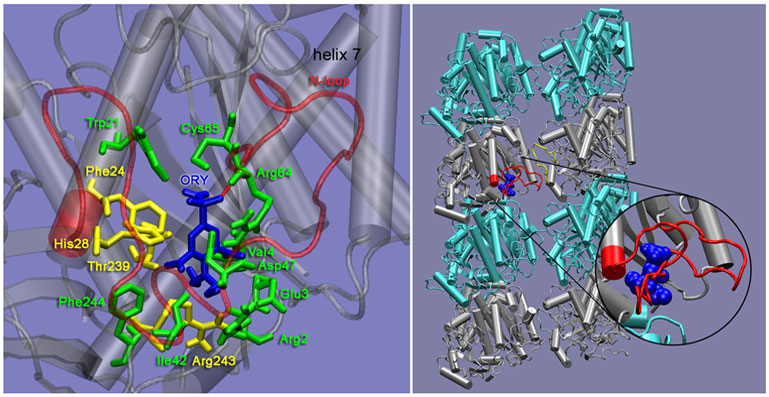
| Figure 11: (Left) A representative structure of oryzalin bound to a-tubulin as predicted by the docking simulations. The binding site includes residues Phe24, His28, Thr239 and Arg243 (yellow) that are mutated in resistant lines. The binding site lies beneath the N loop (translucent red). (Right) Two protofilaments, each consisting of two a-b heterodimers, with bound oryzalin (blue). Oryzalin binds to a-tubulin (silver) beneath the N loop (red); this may interfere with lateral interactions between the N loop and the M loop (yellow) of the adjacent protofilament. | |||
Go to:
Main lab page
Research in our laboratory
Publications